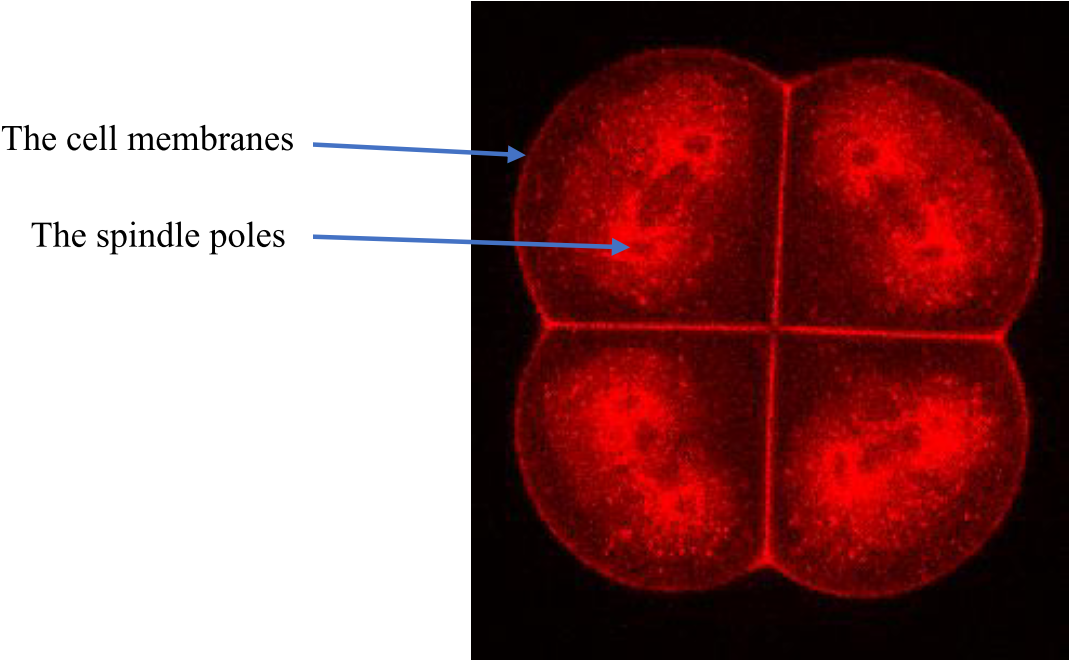
We use the advantages of ascidian early embryogenesis to study the control of 1) Cell number, 2) Cell size, and 3) Cell position and how this 3 aspects are integrated to generate embryo shape.
While the time-dependancy of cell division will set cell number, the spatial aspects of cell division (size and position) rely on complex cell biomechanics and intracellular mechanisms positioning the mitotic spindle. We discovered recently that the timing of cell division informs cell position via cell cycle dependant changes in cell tension and cell adhesion.
Cell Number
We found that cell number is precisely controlled such that all ascidian embryos display a 24-cell stage. This asynchrony is maintained (at the 44-cell stage) and amplified giving rise to a 112-cell gastrula. We have shown in ascidians that cell cycle remodeling depends upon a gene-regulatory network controlled by b-Catenin (McDougall et al, 2012; Dumollard et al., 2013).
Dumollard et al., 2013
Cell size
We studied how unequal cell division (UCD) is controlled in the ascidian embryo. We had previously shown that UCD in Phallusia embryos occurred due to the attraction of one spindle pole towards a cortical structure termed the CAB (for centrosome-attracting body) during prometaphase through anaphase (Prodon et al., 2010, McDougall et al., 2015). We then found that the microtubule depolymerase Kif2 is localized to the CAB and is involved in reducing the size of the proximal aster thereby facilitating the pulling of the spindle pole towards the CAB (Costache et al., 2017). In terms of shaping the whole embryo, CAB ablation completely radializes the embryo indicating that UCD affects the shape of all blastomeres in the early embryo.
Costache et al. 2017
Dumollard et al., 2017
Cell position
Recently we discovered that an apical shape sensing mechanism operates to align all the spindles parallel to the outside (apical) surface of the embryo, orienting them within each cell’s longest axis in the apical plane (with the exception of those oriented by the CAB) (Dumollard et al., 2017). Furthermore relaxation of apical cortical tension during mitosis allows tension oriented cell division reflecting the importance of embryo wide tension/forces in the ascidian embryo (Godard et al 2020).
Intriguingly, tension-oriented cell divisions depend upon cell cycle asynchrony which creates differential cortical tension between soft mitotic and stiff interphasic cells (Godard et al., 2020)
Such conserved cell cycle asynchrony provides a partial cell-biological explanation of the invariant cleavage pattern of ascidian embryos that was elegantly described by Conklin more than a century ago (Conklin, 1905).
Cell position
16-32 cell spindle rotation
Publications
Conklin E.G., (1905) The organization and cell-lineage of the Ascidian Egg. Journal of the Academy of Natural Sciences of Philadelphia 3 (1).
Costache, V., Hebras, C., Pruliere, G. et al. (2017) Kif2 localizes to a subdomain of cortical endoplasmic reticulum that drives asymmetric spindle position. Nat Commun 8, 917 https://doi.org/10.1038/s41467-017-01048-8
Dumollard R, Hebras C, Besnardeau L, McDougall A. (2013) Beta-catenin patterns the cell cycle during maternal-to-zygotic transition in urochordate embryos. Dev Biol.;384(2):331-42.
Dumollard R, Minc N, Salez G, Ben Aicha S, Bekkouche F, Hebras C, Besnardeau L, McDougall A. (2017) The invariant cleavage pattern displayed by ascidian embryos depends on spindle positioning along the cell’s longest axis in the apical plane and relies on asynchronous cell divisions. Elife. Jan 25;6. pii: e19290.
Prodon F, Chenevert J, Hébras C, Dumollard R, Faure E, Gonzalez-Garcia J, Nishida H, Sardet C, McDougall A. (2010) Dual mechanism controls asymmetric spindle position in ascidian germ cell precursors. Development. Jun;137(12):2011-21.
McDougall, A., Chenevert, J. and Dumollard, R. (2012). Cell Cycle Control in Oocytes and during Embryonic Cleavage Cycles in Ascidians. Int Rev Cell Mol Biol. 297, 237-266.
McDougall A, Chenevert J, Pruliere G, Costache V, Hebras C, Salez G, Dumollard R (2015) Centrosomes and spindles in ascidian embryos and eggs. Methods in Cell Biology, Volume 129, 317-39.
Godard B., Dumollard R., Munro E., Chenevert J., Hebras C., McDougall A., Heisenberg C.P.,(2020) Apical Relaxation during Mitotic Rounding Promotes Tension-Oriented Cell Division. Developmental Cell. 55(6):695-706.e4. doi: 10.1016/j.devcel.2020.10.016.
Protocols
Marquage
Embryos
Protocol for labeling the cell membranes and metaphase spindle poles of ascidian embryos
Segmentation manuelle d'image multidimensionnelle
Imaris
Cette segmentation permet d’obtenir à partir d’image multidimensionnelle des objets 3D.
Extraction surface apicale
Imaris
Extraction de la surface apicale d’une cellule préalablement modélisée 3D.
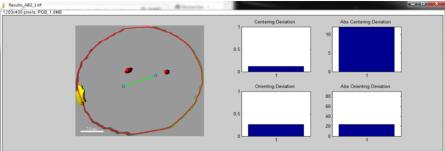
Détermination de l'axe long des blastomères selon le modèle mathématique de Minc
Matlab
Le scripte compare alors l’orientation du fuseau mitotique prédit avec l’orientation du fuseau mitotique observé (« Orienting déviation »).
Il permet également de comparer la position du fuseau mitotique prédit avec la position du fuseau mitotique observé (« Centering déviation »).
Funding
Personnels Enseignants
Carine Barreau
Chercheur | SORBONNE UNIVERSITE
carine.barreau[at]imev-mer.fr
+33 (0) 4 93 76 39 73
Bâtiment Jean-Maetz
Master Biologie Moléculaire et Cellulaire
Parcours Biologie Cellulaire et du Développement & Cellules Souches
DEVELOPPEMENT DES ORGANISMES MARINS (DOMO)
Cette UE se déroule sur 2 semaines et a lieu au laboratoire de Biologie du Développement de Villefranche -sur-Mer (LBDV). Elle inclut l’examen de l’UE d’analyse scientifique (5V089) suivie par les étudiants de la spécialité de Biologie du Développement.
Durant la 1ère semaine, les étudiants participent à des ateliers et des rencontres avec les chercheurs du laboratoire.
Durant la 2ème semaine, les étudiants sont répartis dans les équipes pour y réaliser un mini projet qu’ils présentent le dernier jour du cours. Le cours est donné en anglais pour tout ou partie.
Responsable de l’UE: Carine BARREAU (MCU): carine.barreau [at] obs-vlfr.fr
Master Biologie Intégrative & Physiologie
Parcours Biologie et Bioressources Marines (BBMA)
ORGANISMES MARINS & MODELES BIOLOGIQUES
Cette UE permet aux étudiants de 1ère année de Master de passer 2 semaines à l’Observatoire Océanologique de Villefranche-sur-Mer. Le cours est obligatoire pour les étudiants du Master Biologie Intégrative, parcours Biologie et Bioressources Marines (BBMA) tandis qu’il peut être choisi en option par les étudiants du Master Biologie Moléculaire et Cellulaire (BMC). Les étudiants participent à des ateliers de présentation des organismes marins utilisés par les équipes de recherche du laboratoire (LBDV) et apprennent à les manipuler au cours de travaux pratiques dont les thématiques vont de la Biologie du Développement fondamentale à la toxicologie appliquée.
Responsable de l’UE: Carine BARREAU (MCU): carine.barreau [at] obs-vlfr.fr
Licence de Sciences de la Vie
Parcours Biologie et Bioressources marines
Biologie Des Organismes Marins Et Diversité Des Recherches
Cette UE complémentaire se déroule sur 2 semaines et permet aux étudiants de découvrir les différents aspects (métiers & recherche) de l’Observatoire Océanologique de Villefranche-sur-Mer (OOV). Ateliers et journal clubs sont organisés afin que les étudiants mettent en pratique leurs connaissances théoriques en biologie et développent leur capacité de communication scientifique en français et en anglais.
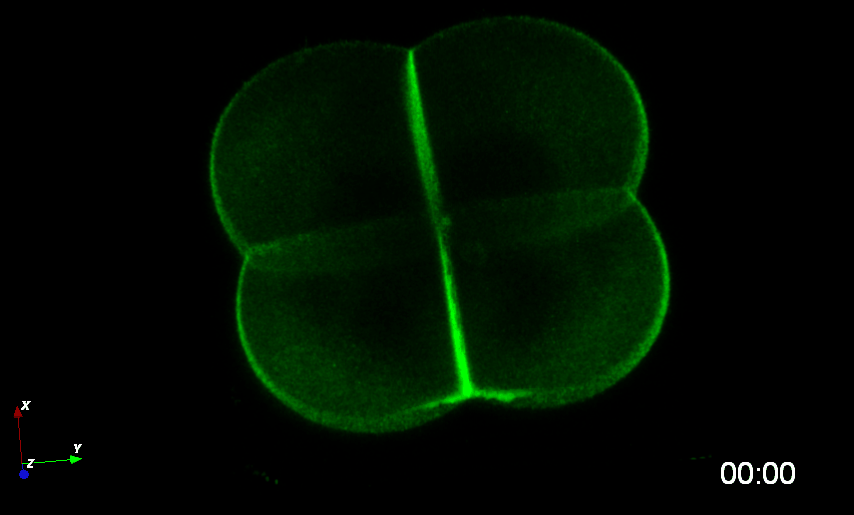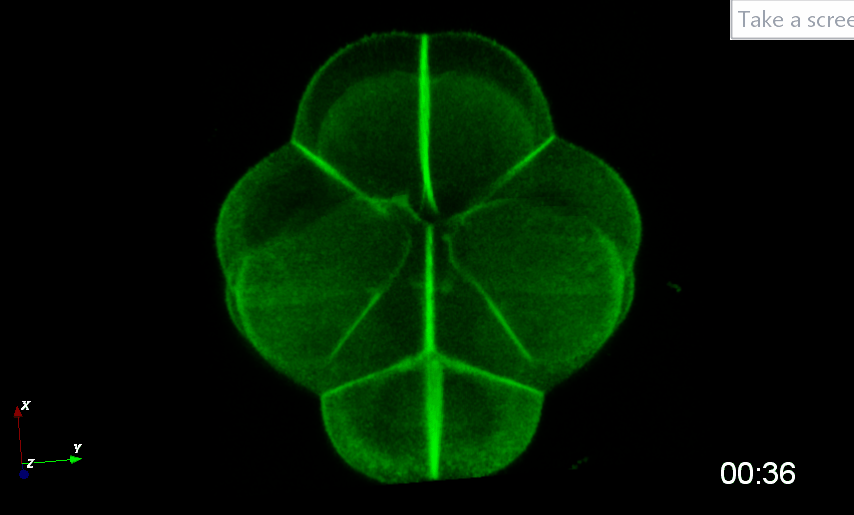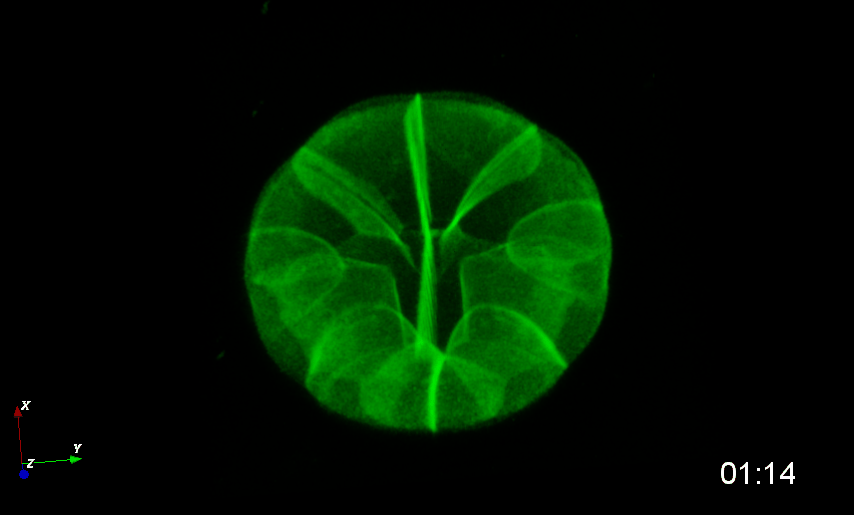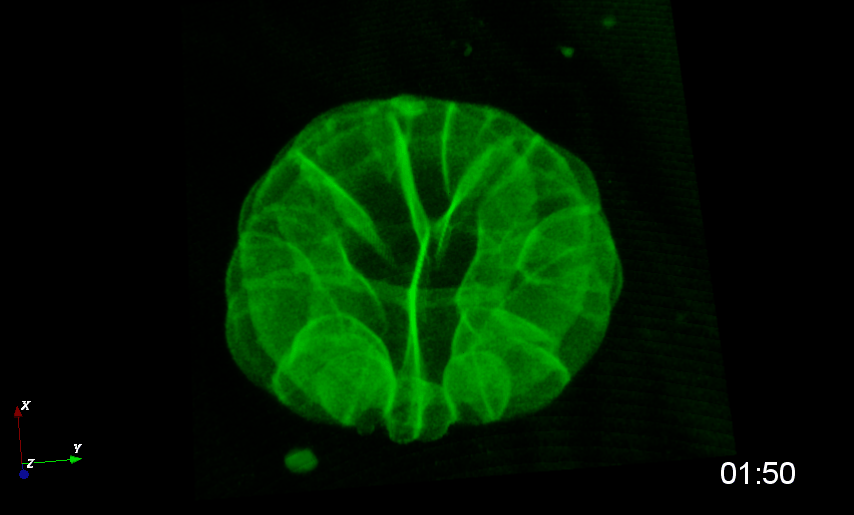
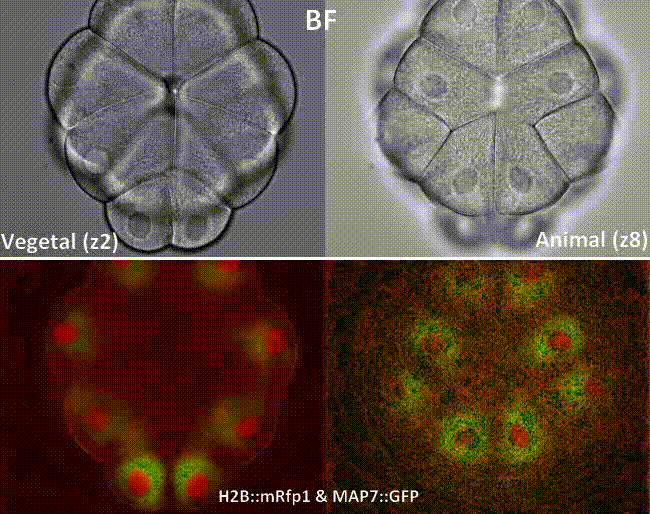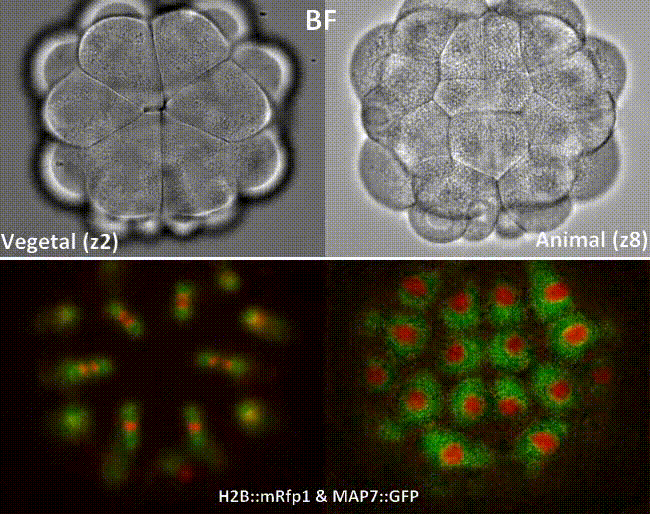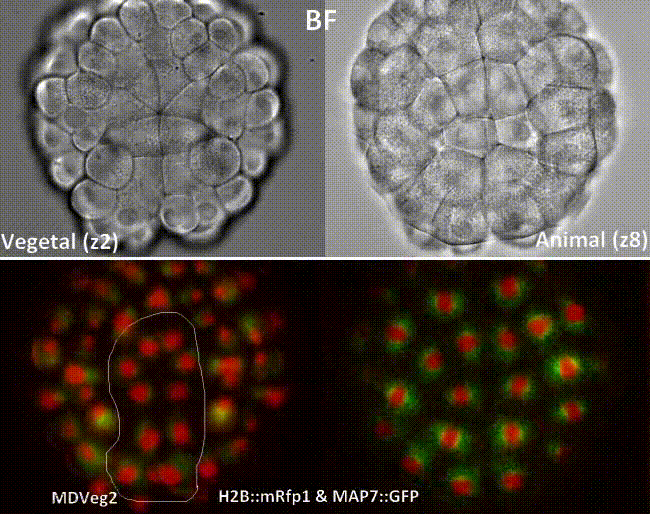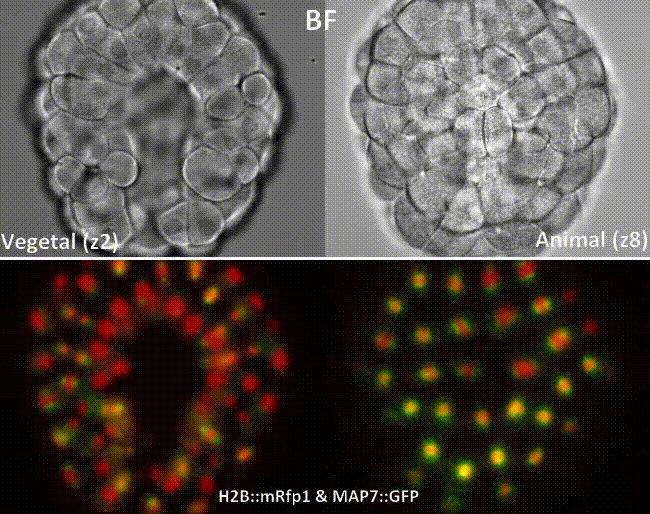 Dumollard et al., 2013
Dumollard et al., 2013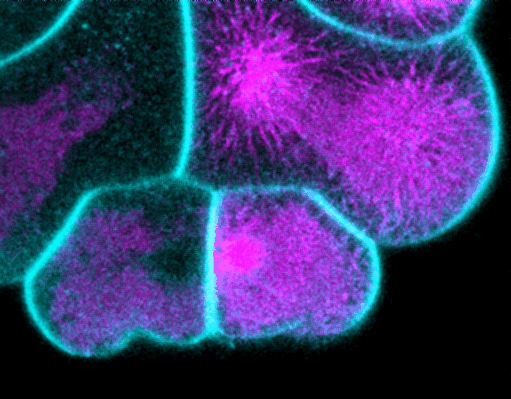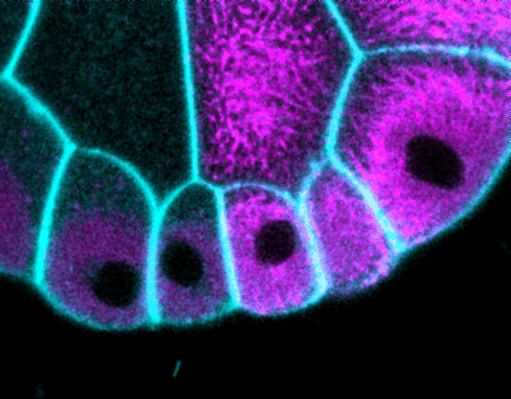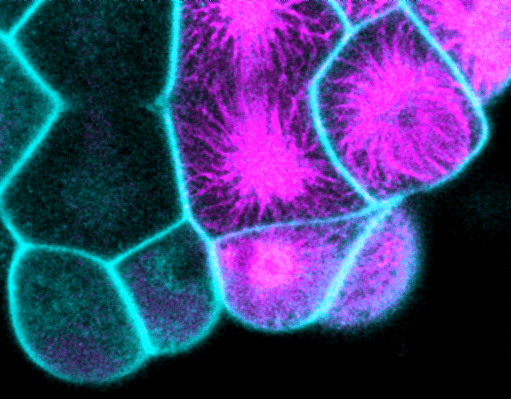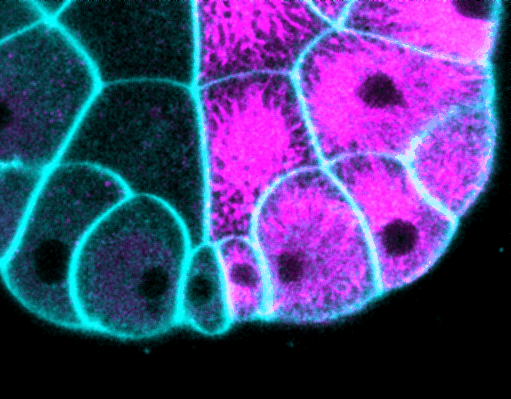 Costache et al. 2017
Costache et al. 2017
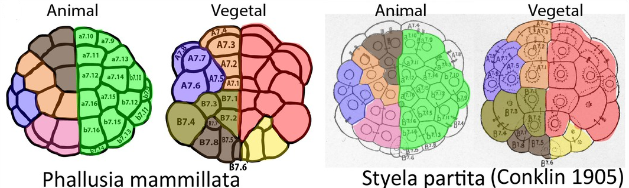
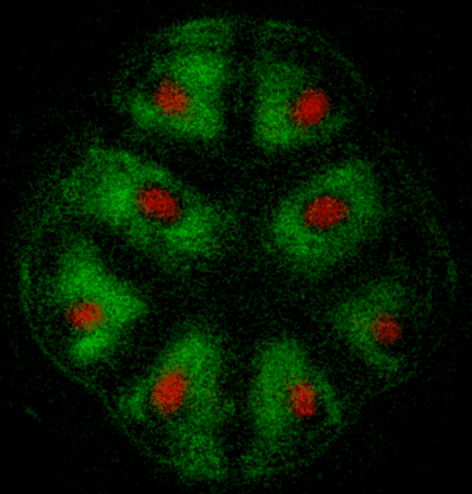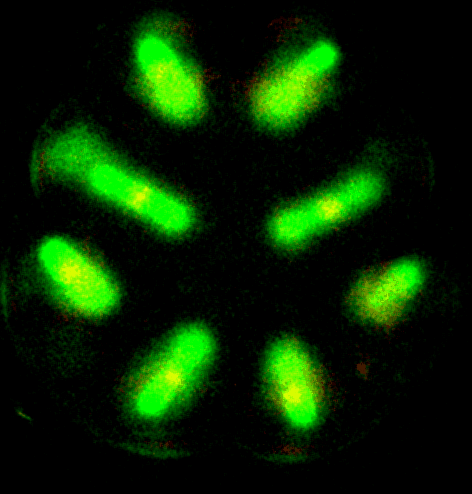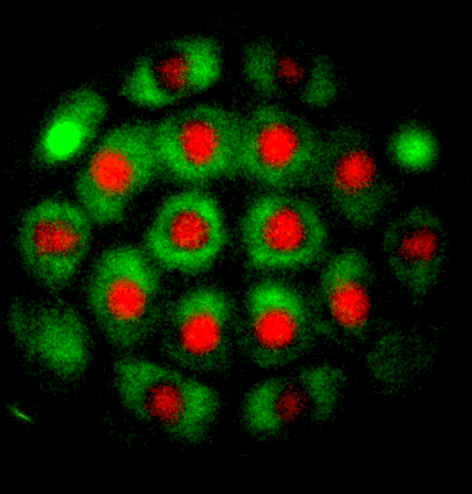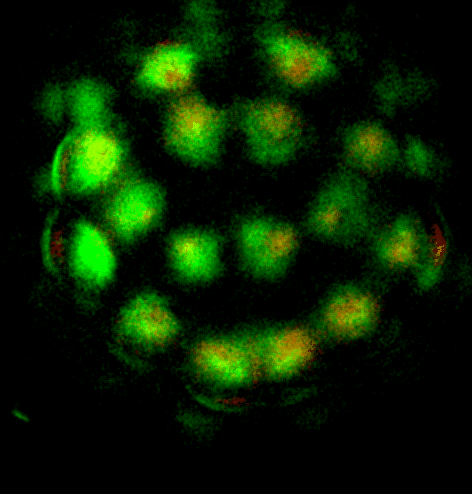 16-32 cell spindle rotation
16-32 cell spindle rotation